Channels


We just wrapped collecting 10,000+ hours of on-demand egocentric human activity data for a major robotics company. Are there any specific tools that you can recommend to data annotation. Happy to share our sample dataset with anyone working on manipulation or foundation models! submitted by /u/RemarkableFold888 [link] [Kommentare]
A story exploring the serendipitious effects of a world in which
A conjectural theory of cognitive inequality
In 2022, I wrote about the damning fall of events tech company Pollen, founded by Callum Negus-Fancey. The short of it: Pollen seemed to have pulled off the improbable feat of building a business in the notoriously low margin industry of events, surviving Covid-19, and building a solid

About the author A human voice behind WNF Nicolás Halaban is an Argentine writer and AI-assisted creator exploring the meeting point between Scripture, language, technology, and spiritual imagination. The Word, The Name, The Fire was born from a long dialogue with artificial intelligence — not as doctrine, but as a...

Chinese scientists have discovered a way to combine high-tech light-absorbing nano-technology with more conventional water-based carbon pigment spray to coat car bodies with a durable ultra-black finish. The result is a black that creates a ‘black-hole’ effect of virtually no reflected light at all.

Building a custom octocopter from scratch with an RL controller that sustains flight through motor failures. Follow the build log.
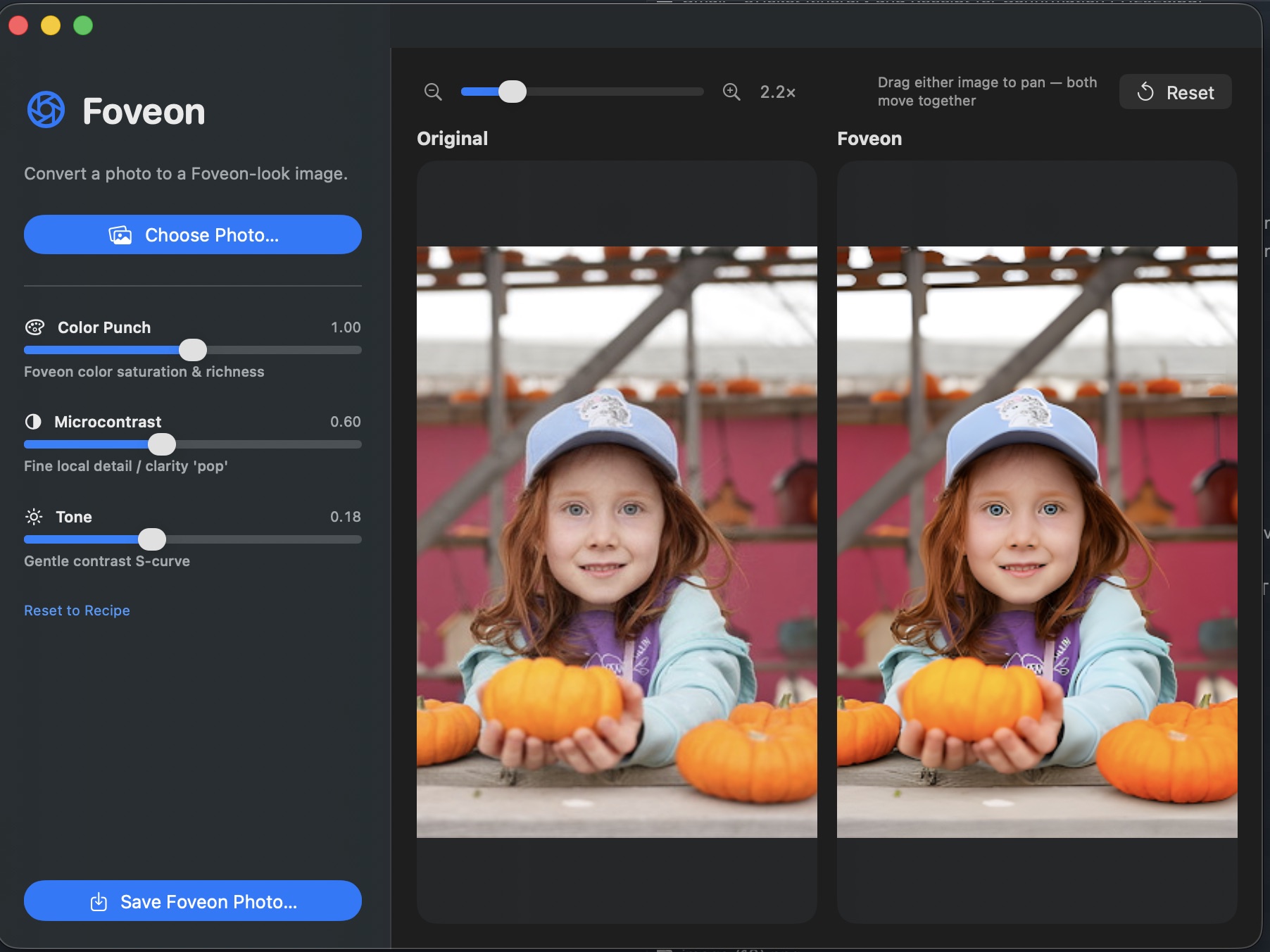
Foveon A neural sensor translator. Takes a photo from any Bayer-array camera and renders it as if it were shot on a Sigma DP2 Merrill — the Foveon X3 stacked-sensor look, with the colour and microdetail Foveon is famous for, on hardware you already own. Under the hood: a modified U-Net with an extra layer injected between the encoder bottleneck and the upsampling decoder. The injected channel carries a one-dimensional encoding of three-layer pixel-stack structure — the B·G·R photodiode column that a Foveon sensor captures and a Bayer sensor can’t. Trained end-to-end against matched Bayer → Merrill scene pairs. U-Net+1D Modified U-Netwith 3-layer pixelinjection at bottleneck Bayer → X3 Bayer CFA in,Foveon X3 stacked-sensor look out DP2 Merrill Trained on matchedscene pairs againstSigma DP2 Merrill ⤓ Download Foveon.dmg macOS 13+ · Apple Silicon 33 MB · signed DMG installer · unverified-developer gatekeeper: right-click Open the first time Foveon — macOS app. Choose a photo, drag the sliders, save the result. What it is Most digital cameras capture colour through a Bayer colour filter array: each photosite sees only one of R, G, or B, and the other two channels are interpolated from the neighbours (demosaiced). It’s efficient, but it costs you. The interpolation introduces colour fringing on sharp edges, smears fine detail, and produces the “digital” micro-contrast that even high-end Bayer cameras can’t fully shake. The Foveon X3 sensor — Sigma’s now-rare design used in the DP1, DP2, and DP3 Merrill cameras — works the way colour film does. Three photodiode layers are stacked vertically at every single pixel position. The top layer absorbs blue, the middle layer green, the bottom layer red. Every pixel captures the full colour. No interpolation, no demosaicing artefacts, no false detail. The result is the “Foveon look”: extraordinary microdetail and a particular colour rendition — warm, dimensional, almost slide-film — that people build entire camera systems around. Foveon (the app) is a neural network that learns the mapping between the two. Feed it a JPEG or RAW from a normal Bayer camera (phone, mirrorless, DSLR) and it predicts what the same scene would look like shot on a Foveon X3 sensor. Geometry stays the same; colour, tonality, and micro-detail rendering shift toward the Merrill side of the training distribution. Bayer vs Foveon — the structural problem Bayer CFA One colour per pixel R G ×2 B Each photosite captures exactly one colour. The other two channels are guessed from the neighbours. The guess is what creates the “digital” signature. Foveon X3 Three layers per pixel red (bottom) green (middle) blue (top) stacked photodiodes, one per pixel Every pixel records R, G, and B separately at the same location. No interpolation. No false colour. The dimensional quality Merrill shooters chase. The architecture The core is a standard convolutional U-Net: an encoder that downsamples the input image into a compact feature bottleneck, paired with a decoder that upsamples back to full resolution, with skip connections at every level so fine spatial detail survives the trip through the bottleneck. The modification is a single new layer dropped in between the encoder’s final downsampling block and the decoder’s first upsampling block: a 1D pixel-stack injection layer that concatenates a one-dimensional encoding of how colour absorbs through silicon depth on a real Foveon sensor — blue first, then green, then red. The decoder learns to use this prior to reconstruct the kind of inter-channel coupling that real X3 captures exhibit — chroma that’s registered with luminance instead of interpolated against it. U-Net architecture diagram. Encoder ENC1 (C=64) → ENC2 (C=128) → ENC3 (C=256) → ENC4 (C=512) → Bottleneck (C=1024) → 1D Pixel-Stack Injection (B·G·R depth prior) → DEC4 (C=512) → DEC3 (C=256) → DEC2 (C=128) → DEC1 (C=64) → output, with skip connections from each encoder level to the matching decoder level. Encoder (blue) downsamples the Bayer input. The 1D injection layer (orange) concatenates the Foveon B·G·R depth prior at the bottleneck. Decoder (purple) upsamples back to full resolution. Skip connections (dashed) carry pre-bottleneck spatial detail across to the matching decoder level — standard U-Net, drawn here for completeness. The novel piece is the orange block. Why inject at the bottleneck The encoder has just stripped spatial resolution to focus on semantic content; the decoder is about to reconstruct it back. That’s exactly the moment to inject the prior that says “reconstruct as if the sensor were stacked, not mosaiced.” Inject earlier and the encoder learns to ignore it; inject later and the decoder has already committed to a demosaic-style chroma reconstruction. Why one-dimensional The injection encodes the order in which colour absorbs through silicon depth on a real Foveon sensor — blue at the top, green in the middle, red at the bottom. That ordering is a 1D signal per pixel column, not a 2D spatial structure. Keeping it 1D keeps the parameter count low and forces the network to learn the inter-channel coupling rather than memorise a per-region lookup. Training The model is trained end-to-end against matched scene pairs: the same subject captured simultaneously by a Bayer-sensor camera and a Sigma DP2 Merrill, the two images aligned and cropped to identical framing. The Bayer image is the network input; the Merrill image is the reconstruction target. STEP 1 Capture matched pairs A Bayer-sensor camera and a Sigma DP2 Merrill are tripod-mounted side-by-side. Each scene is photographed simultaneously by both. We bias capture toward the categories where Foveon rendering differs most visibly from Bayer: outdoor light, foliage, skin tones, fabric, water, metallic surfaces. STEP 2 Align & crop For each pair we run a homography-based alignment (feature-matched on luminance) so the two images share an identical pixel grid despite the different sensor sizes, focal lengths, and viewpoints. The aligned tiles are cropped to common framing and resampled to a shared resolution. Misaligned pairs are filtered out. STEP 3 Develop the Merrill target DP2 Merrill RAW (.X3F) files are developed in Sigma Photo Pro with neutral parameters — zero added sharpening, native colour profile, no creative tone curve. This is the “ground truth” the network is asked to predict: what the X3 sensor saw, minimally interpreted. STEP 4 Tile & augment Each aligned pair is sliced into overlapping 256×256 tiles for training (the U-Net handles arbitrary input sizes at inference, but training on fixed tiles keeps batches efficient). Augmentations: random crop, horizontal flip, small rotation, exposure jitter on both halves of the pair in lock-step. STEP 5 Loss composition Three components, summed with weights tuned to keep colour and detail in balance: L1 pixel loss anchors absolute colour fidelity; VGG perceptual loss carries microdetail and texture (the network learns the Merrill look, not just the average pixel value); a small TV smoothness penalty keeps the output from inventing sharpening artefacts in flat regions. STEP 6 Train & validate AdamW optimiser at lr = 2e-4, cosine schedule, mixed-precision on a single 24 GB GPU. Batch size 16 tiles. We hold out roughly 5% of pairs as a validation set stratified by category (skin, foliage, fabric, etc.) so the model isn’t accidentally memorising a few scenes. Best checkpoint is the one that minimises validation perceptual loss, not L1 — perceptual loss tracks the X3 look more reliably than pixel error. What it produces A schematic comparison: what the same uniform skin patch looks like coming out of a Bayer demosaic pipeline (left) versus what the network predicts the Merrill would have rendered (right). The Bayer side has slight chroma drift between cells; the Merrill side is uniform. INPUT Bayer demosaic visible chroma noisebetween adjacent pixels —the demosaic signature OUTPUT Foveon → X3 look uniform chroma per region,microdetail preserved —the Foveon X3 signature The app Foveon ships as a native macOS app with a clean three-pane workflow: pick a photo on the left, watch the preview update on the right. Three sliders shape the conversion: Color Punch Controls how strongly the Foveon colour rendition is applied. At 0 the output sits closer to the original Bayer colours; at 1.0 (default) the full Merrill warmth and saturation come through. Microcontrast The Foveon look isn’t about sharpening — it’s about fine local detail clarity. This slider lifts that quality on top of the network output without introducing the halo artefacts that conventional unsharp masking would. Tone A gentle contrast S-curve in the output highlights and shadows. Default 0.18 matches Sigma Photo Pro’s neutral tone; push it for a more cinematic look, drop it for a flatter starting point you can grade externally. Drag either preview panel to pan; both panels move together so the same crop is always in view on both sides. Reset snaps the view back to fit, Save Foveon Photo… writes the result. Install & use Foveon runs natively on Apple Silicon. A 24-megapixel input converts in well under a second on an M2; the model uses MPS for inference and stays within ~3 GB of unified memory. ⤓ Download Foveon.dmg macOS 13+ · Apple Silicon · 33 MB Open the DMG and drag Foveon.app into Applications. First launch: right-click the app and choose Open (the DMG is signed but not notarised by Apple yet, so Gatekeeper asks once).

PAC-832 is the first drug of its kind — it works by selectively blocking a receptor called “galanin receptor 1,” or GalR1. The drug has sub-micromolar potency for GalR1 and >30x selectivity over GalR2/3. When administered to mice, it significantly improves their memory across multiple different memory tests. PAC-832 also has excellent manufacturability, low toxicity, and great pharmacokinetics, including passing the blood-brain barrier — all critical things needed for a neurological drug to succeed. Motivation behind PAC-832 The primary symptom of Alzheimer’s disease (AD) — memory loss — has long been hypothesized to arise from the loss of neurons that produce the neurotransmitter acetylcholine (ACh) in certain important brain regions, primarily the basal forebrain. This is known as the “cholinergic hypothesis of AD,” which forms the basis for how the AD drug donepezil works. Donepezil increases the amount of ACh between neurons by blocking an enzyme called acetylcholinesterase (which is responsible for breaking down ACh). In doing so, it’s been shown to reverse memory loss in AD patients. Acetylcholine (ACh) Donepezil Donepezil is far from perfect and was discovered way back in 1983, yet shockingly the entire AD drug development field has been unable to improve on it after 40+ years of effort. During this time, the AD field has mostly focused on developing drugs that target amyloid-beta plaques and other “disease-modifying” mechanisms. This effort produced several new FDA-approved drugs in recent years which reduce plaque levels (like lecanemab and donanemab), yet none of these drugs meaningfully slow AD disease progression or outperform donepezil for managing AD symptoms, and they carry significant new safety risks like increased incidence of brain hemorrhage/stroke. Because of this, doctors are unwilling to prescribe these new drugs. Almost nobody uses them. Meanwhile, donepezil has remained the most effective, safe, and widely-used AD drug from its discovery up to today. Given the underwhelming performance of anti-amyloid drugs, it’s a good time to focus on alternative AD drug mechanisms. I believe that the optimal place to look is the cholinergic pathway, which unlike more contemporary AD targets has a large body of the best possible supporting evidence — decades of real, human clinical data from cholinesterase inhibitors. This line of thinking led me to develop PAC-832. Like donepezil and related cholinesterase inhibitors (galantamine, rivastigmine), PAC-832 ultimately increases ACh levels in the brain. However, PAC-832 is not structurally related to any existing cholinesterase inhibitors, and it doesn’t directly target the cholinergic pathway. Instead, it works through a new mechanism revolving around this complex, underappreciated molecule called “galanin.” A brief history of the galanin-AD hypothesis Galanin is a 30-amino acid neuropeptide — a small protein that acts as a signaling molecule in the brain. It was discovered in 1978. Though less well known than dopamine or serotonin, galanin is widely produced by neurons throughout the central and peripheral nervous system. Galanin signaling has been shown to regulate pain, metabolism, mood, sleep, and memory. It lurks in the background, quietly influencing many of our physiological functions on a daily basis. 3D structure of human galanin Galanin dysregulation was first tied to AD pathology in the 1980s, 10 years after galanin’s discovery. Chan-Palay, Beal et al., and others observed that the basal forebrain from deceased AD patients contained many more galanin-producing neurons and higher galanin expression than healthy brains. Meanwhile, Fisone et al. and Dutar et al. observed that galanin treatment onto slices from rat and monkey brains prevented the neurons from releasing ACh. At the behavioral level, impaired memory was observed by McDonald et al. in rodents with galanin injected into their brains. Steiner et al. observed similar results in rodents genetically engineered to overexpress galanin. Along with other supporting studies, these observations outlined a mechanism where galanin contributed to memory loss in AD by preventing ACh release in the basal forebrain, though neither galanin’s molecular mechanism nor the cause of galanin overexpression was understood at the time. This in turn led people in the 90s to hypothesize that a drug that ‘blocked’ the action of galanin could increase ACh levels and improve cognitive function in AD patients, similar to how cholinesterase inhibitors like donepezil work. Source: McDonald et al. 1998, J Neurosci In the 2000s, the galanin-AD hypothesis was complicated by the emergence of data showing that galanin had neuroprotective effects in certain contexts. O’Meara et al. showed that galanin knock-out mice had significantly fewer cholinergic neurons in the basal forebrain, demonstrating that galanin was necessary for normal neuronal development. Meanwhile, Ding et al. observed that galanin protected cholinergic neurons from death caused by amyloid beta plaques. These findings, along with related reports, provided a surprising explanation for why cholinergic neurons in AD patients contained so much galanin in the first place — it was likely the brain trying to protect the neurons from neurotoxic factors brought on by AD progression, and the suppression of ACh release was collateral damage. This in turn suggested that a blanket inhibitor of galanin across all contexts ran the risk of causing harm by blocking galanin’s neuroprotective effects. Concurrent molecular research helped resolve the seemingly contradictory neuroinhibitory vs neuroprotective effects of galanin. By the 2000s, it was known that galanin activated three different receptors, which were named galanin receptor 1, 2, and 3 (GalR1–3 for short). GalR1 and GalR3 were found to activate the “Gi/o signaling pathway,” a powerful molecular brake that suppresses neurotransmitter release. This signaling is desirable in certain non-AD contexts — for example, in overactive neurons prone to causing seizures. However, in cholinergic neurons relevant for memory, decreased ACh release is detrimental. This finding explains how galanin overexpression leads to ACh suppression and memory loss on a molecular level — via activating GalR1/3 and the Gi/o pathway. Schematic of galanin signaling pathways mediated by GalR1/2/3.Source: Lang et al. 2007, Pharmacology & Therapeutics On the other hand, GalR2 was found to activate a different pathway — the Gq/11 pathway, which broadly results in cell activation; the opposite of Gi/o signaling. Moreover, various studies (Mahoney et al., Elliot-Hunt et al., and others) found that the neuroprotective effects of galanin were downstream of this pathway and mediated through GalR2. A viable path for a pharmacological inhibitor of galanin emerged from these findings. Rather than blocking galanin activity across all contexts (which would also block its protective effects), one could in theory design an inhibitor that selectively blocks ONLY GalR1 and/or GalR3 (thus removing the brake on ACh release), while having no effect on GalR2 (thus preserving the protective Gq/11 signaling). Between GalR1 and GalR3, GalR1 was the more attractive target due to its higher expression in the brain. Source: Hokfelt and Tatemoto 2010, Galanin Previous galanin antagonist drug development At the same time that researchers were working out galanin’s biology in the 90s and onward, a parallel effort was underway to develop drugs that modulate galanin activity, with the hopes of finding a treatment/cure for any of the diseases tied to galanin dysregulation. The initial batch of GalR antagonists/agonists developed in the 90s were all peptides. They were neither receptor subtype-specific nor able to pass the blood-brain barrier due to their size. They had to be administered by direct injection into the brain — feasible for a mouse, obviously not feasible for a human. Still, these early drugs provided important proof of concept data for galanin antagonist drugs. For example, McDonald and Crawley found that the peptide antagonist M40 (a galanin analog) was able to reverse cognitive deficits caused by galanin in rats. The first small molecule inhibitors of GalR1 with potential to pass the blood-brain barrier appeared in the early 2000s. A few high-throughput screens were carried out for GalR1 binding, producing two promising lead compounds/series with good affinity for GalR1 (Chu et al. 1997, Scott et al. 2000). However, none of these compounds were viable for further development. One of them, Sch 202596, was a fungal metabolite that was too difficult to synthesize and modify. The other, the dithiin/dithiepine-1,1,4,4-tetroxide series, suffered from intractable “reactivity and solubility issues.” Sch 202596Chu et al. 1997, Tetrahedron Lett. Dithiepine-1,1,4,4-tetroxide seriesScott et al. 2000, Bioorg. Med. Chem. Since then, there has been no further development of small molecule GalR1 antagonists — partially due to the practical challenges like the ones listed, and partially due to the fact that the galanin-AD hypothesis fell out of fashion in favor of other AD mechanisms. As of today, there are still no FDA-approved drugs that target galanin signaling. The closest thing so far has been the GalR3-selective small molecule antagonist HT-2157, which entered clinical trials in 2011 for depression. However, the drug did not progress beyond Phase 1 due to safety issues. HT-2157 Enter PAC-832 To summarize everything so far: there is a large body of basic research dating back to the 1980s that supports galanin signaling as a promising target for treating cognitive decline in AD. However, various issues have prevented the development of a successful galanin antagonist drug, including (1) messy galanin biology that took decades to elucidate, which requires subtype-specific receptor antagonism that avoids GalR2, (2) poor chemical properties for the few small molecule GalR1 antagonists that were actually discovered, and (3) a lack of precedent from existing successful galanin-targeting drugs in the clinic. PAC-832 solves issues (1) and (2), and will hopefully be the first drug to solve (3). PAC-832 mechanism of action PAC-832 selectively inhibits only GalR1 and not GalR2/3. It has sub-micromolar potency toward GalR1 (IC50 = 0.28 μM) in an in vitro GalR1 activity assay, while showing no detectable activity at GalR2 or GalR3 at concentrations up to 10 μM. Thus, PAC-832 fits the desired profile of a galanin antagonist — potent inhibition of GalR1 to release the brake on ACh release, while having no effect on GalR2 to preserve neuroprotective signaling pathways. The expected downstream effects of GalR1 inhibition are (1) increased ACh release, and (2) improved cognition. We confirmed that PAC-832 produces each of these observations in separate experiments. In an in vitro ACh release assay, PAC-832 concentration-dependently rescues ACh levels when co-administered with galanin, confirming (1). Meanwhile, PAC-832 dose-dependently improves memory in two rodent behavioral assays, with comparable efficacy to donepezil, confirming (2). Last but certainly not least, PAC-832 exhibits excellent developability. It has good stability, solubility, and ease of synthesis. It also has great pharmacokinetics, with 90% oral bioavailability and a brain-to-plasma ratio (Kp) of 0.7, as well as low toxicity (well-tolerated at doses up to 1000 mg/kg in mice). I’m now preparing PAC-832 for Phase 1 clinical trials, which should be started within a year. PAC-832 will be the first GalR1 antagonist to reach the clinic.
AICoS - An AI Chief of Staff platform that every CEO will ever need. - suhasbhairav/ai-chief-of-staff
